![]()
MINISTERIO DE EDUCACIÓN Y CULTURA
Secretaría de Estado de Universidades, Investigación y Desarrollo
Dirección General de Enseñanza Superior e Investigación Científica
|
|
MINISTERIO DE EDUCACIÓN Y CULTURA Secretaría de Estado de Universidades, Investigación y Desarrollo Dirección General de Enseñanza Superior e Investigación Científica |
Solicitud de Ayuda para Proyectos de
Investigación y Desarrollo Tecnológico
PROGRAMA NACIONAL EN EL QUE SE ENCUADRA LA SOLICITUD
CIENCIA Y TECNOLOGÍA MARINA (CYTMAR)
MODALIDAD A LA QUE SE PRESENTA LA SOLICITUD: ý
A o BPROYECTO COORDINADO
NÚMERO DE SUBPROYECTOS DE QUE CONSTA
3
INDIQUE EL ÁREA O ÁREAS DE LA ANEP EN LAS QUE CONSIDERA DEBE EVALUARSE LA SOLICITUD:
|
o Física y Matemáticas o Química ý Biología de Organismos y Sistemas
o Ciencias de la Tierra y del Espacio o Ciencias Sociales o Ciencias Económicas y Jurídicas o Ciencias Humanas o Agricultura |
o Ganadería o Industria o Medicina o Tecnología de Alimentos o Tecnología de la Información y las Comunicaciones o Tecnología de Materiales o Tecnología del Medio Ambiente o Tecnología Mecánica y Textil o Tecnología Química |
DOCUMENTACIÓN QUE SE ADJUNTA:
ý
Firma de conformidad del representante legal del organismo solicitanteý
Memoriaý
Relación de personal investigadorý
Curriculum vitae, firma de conformidad y copia DNI del personal investigadorý Original y copias de la documentación
COMISIÓN INTERMINISTERIAL DE CIENCIA Y TECNOLOGIA
PROYECTO COORDINADO
DATOS DEL PROYECTO
1. Título:
2. Objetivo científico-técnico: 2.2
3. Clasificación UNESCO: 251001
4. Duración (en años): 3
5. Nº de investigadores: 12
DATOS DE LA INSTITUCIÓN SOLICITANTE
6. Organismo: UNIVERSIDAD DE VIGO
7. C.I.F.: Q-8650002-B
DATOS DEL INVESTIGADOR RESPONSABLE
8. Apellidos: FERNÁNDEZ SUÁREZ
9. Nombre: EMILIO MANUEL
10. Centro: FACULTAD DE CIENCIAS
11. Departamento: ECOLOGÍA Y BIOLOGÍA ANIMAL
12. Teléfono (prefijo, número, extensión): 986-812591
13. Telefax: 986-812556
14. Correo electrónico: [email protected]
15. Dirección postal completa: FACULTAD DE CIENCIAS. UNIVERSIDAD DE VIGO. CAMPUS LAGOAS-MARCOSENDE. 36200 VIGO.
16.
DATOS AYUDA SOLICITADA (EN MILES DE PTAS.)|
1ª Anualidad |
2ª Anualidad |
3ª Anualidad |
Total |
|
Personal |
1620 |
1620 |
1620 |
4860 |
Material inventariable |
3586 |
0 |
0 |
3586 |
Material fungible |
2911 |
4496 |
2136 |
9543 |
Viajes y dietas |
2735 |
5069 |
790 |
8595 |
Otros gastos |
965 |
1265 |
420 |
2650 |
Total solicitado |
118178 |
12450 |
4966 |
29234 |
Costes indirectos (12%) |
1418 |
1494 |
596 |
3508 |
Total a librar a la entidad |
13236 |
13944 |
5562 |
32742 |
Conforme la Autoridad que representa legalmente al Organismo solicitante, que declara conocer las normas de la convocatoria y, en caso de ser financiada la solicitud, autoriza, a efectos de lo previsto en la Ley Orgánica 5/1992, de 29 de octubre, la utilización de la información contenida en la misma para su difusión en bases de datos de I+D:
D. Salustiano Mato de la Iglesia
Cargo: Vicerrector de Investigación Firma y sello
_____de___________________de 1999
Ilmo. Sr. Director General de ENSEÑANZA SUPERIOR E INVESTIGACIÓN CIENTÍFICA.
C/ Rosario Pino, 14-16, planta 7ª. 28020 MADRID
COMISIÓN INTERMINISTERIAL DE CIENCIA Y TECNOLOGÍA
MEMORIA
INVESTIGADOR RESPONSABLE O COORDINADOR DEL PROYECTO:
EMILIO M. FERNÁNDEZ SUÁREZ
TITULO: CIRculación de CArbono y Nitrógeno en el océano Atlántico (CIRCANA)
PALABRAS CLAVE (Breve descripción, por medio de palabras clave, de la temática, tecnologías empleadas y aplicaciones del proyecto):
Ciclos biogeoquímicos marinos, carbono, nitrógeno, plancton, producción primaria, respiración, reciclado de nutrientes, variabilidad espacial a gran escala, Océano Atlántico.
RESUMEN (con clara exposición de los objetivos y potenciales aplicaciones del proyecto)
Para determinar el papel del plancton marino en la regulación de los ciclos biogeoquímicos de la biosfera es preciso utilizar aproximaciones multidisciplinares que consideren escalas espaciales globales. El proyecto CIRCANA tiene como objetivo la cuantificación de los flujos de carbono y nitrógeno asociados a los tipos de organización trófica planctónica característicos de las diferentes regiones del Océano Atlántico entre 50°N y 50°S. El trabajo propuesto aprovecha la oportunidad brindada por el programa británico Atlantic Meridional Transect (AMT) para realizar tres campañas oceanográficas entre el Reino Unido y las Islas Malvinas. A lo largo de estos transectos, se determinarán 1) la estructura de biomasa del plancton; 2) la importancia relativa de cada clase de tamaño de productores primarios; 3) la tasa de producción de carbono orgánico disuelto y su tasa de utilización por las bacterias; 4) la contribución del zooplancton al metabolismo del ecosistema planctónico; 5) la relación entre producción nueva y regenerada considerando los componentes particulado y disuelto y 6) el balance neto entre producción y respiración en la capa superficial del océano. Los resultados obtenidos permitirán calcular balances tróficos sinópticos para cada provincia biogeoquímica y evaluar el efecto neto de éstas sobre el reparto de CO2 entre el océano y la atmósfera.
TITLE:
Carbon and Nitrogen Cycling in the Atlantic OceanKEY WORDS:
Marine biogeochemical cycles, carbon, nitrogen, plankton, primary production, respiration, nutrient recycling, large-scale spatial varibility, Atlantic Ocean.SUMMARY
A multidisciplinary approach over global spatial scales is required if we are to determine the role of marine plankton for the regulation of biogeochemical cycles in the biosphere. The main objective of CIRCANA is to quantify the carbon and nitrogen fluxes associated with each of the planktonic trophic structures encountered in the Atlantic Ocean from 50°N to 50°S. The proposed work takes advantage of the UK-funded Atlantic Meridional Transect (AMT) programme in order to conduct three oceanographic cruises between UK and Falkland Islands. The following variables will be determined along each transect: 1) plankton biomass structure; 2) relative importance of primary producers of each size class; 3) production rate of dissolved organic carbon and its rate of consumption by bacteria; 4) zooplankton contribution to total planktonic metabolism; 5) new to regenerated production ratio when considering both particulate and dissolved components and 6) net balance between production and respiration in the upper layers of the ocean. The results obtained will be used to calculate synoptically a trophic budget for each biogeochemical province, thus allowing an assessment of their net effect on the CO2 partitioning between the ocean and the atmosphere.
INTRODUCCIÓN
Antecedentes y estado actual de los aspectos científico-técnicos, incluyendo la bibliografía más relevante.
Indíquense también posibles coincidencias con actividades de otros grupos o entidades públicas y privadas en España, así como la posible vinculación del tema propuesto con temáticas semejantes desarrolladas en ámbitos internacionales, preferentemente europeos.
En la Modalidad A, destaque la originalidad y el grado de innovación del tema propuesto.
En la Modalidad B, destaque la oportunidad del tema propuesto, teniendo en cuenta la posibilidad de transferencia de resultados a sectores empresariales o sociales.
El océano en el contexto del cambio global
Cálculos recientes basados en la integración de modelos de producción primaria marina y terrestre indican que la fijación de carbono (C) en los océanos (estimada en 48.5 GtC año-1) representa anualmente el 46% de la producción global de la Biosfera (Field et al., 1998).
La producción biológica de carbono orgánico en la capa eufótica del océano y su posterior remineralización en aguas profundas es responsable de tres cuartas partes del gradiente vertical de S CO2 disuelto en el agua (Volk y Hoffert, 1985). La existencia de este gradiente hace que el océano se comporte globalmente como un sumidero de C antropogénico, retirando ~ 2 GtC de las ~ 7 GtC que son emitidas anualmente a la atmósfera a consecuencia de las actividades humanas (Sarmiento y Bender, 1994). En escalas temporales geológicas, la profunda influencia de la biología de los océanos sobre el ciclo global del C queda de manifiesto en la fuerte correlación inversa que existe entre la productividad marina y los niveles atmósfericos de CO2 durante los últimos 160 000 años (Williamson y Holligan, 1990).
Es improbable que en escalas cortas (100-102 años) se produzca un cambio en la cantidad de C presente en el compartimento planctónico, sobre todo teniendo en cuenta su relativamente alta tasa de renovación. Sin embargo, sí son posibles varios mecanismos de cambio que afectan al flujo de CO2 entre el océano y la atmósfera a través de modificaciones en las tasas de producción primaria y, consecuentemente, en los patrones de circulación de materia por la red trófica planctónica. Entre estos mecanismos cabe mencionar tres:
La importancia de las regiones oligotróficas
En la actualidad aún no es posible realizar predicciones cuantitativas sobre la interacción de todos los mecanismos de retroalimentación que existen entre el funcionamiento de los ecosistemas marinos y los procesos climáticos y geofísicos que afectan a la atmósfera y al océano. Sin embargo, se sabe ya que los procesos biológicos del océano son suficientemente sensibles al forzamiento físico como para desempeñar un papel importante en la regulación del clima en escalas intermedias, generando cambios en la partición de CO2 entre el océano y la atmósfera o amplificando los cambios ya existentes (Falkowski et al., 1998).
La adquisición de una capacidad predictiva mínima en este campo pasa necesariamente por alcanzar una mayor comprensión de los patrones de circulación de materia a través de los ecosistemas marinos. En las últimas décadas se han investigado intensamente los ciclos de carbono, nitrógeno, fósforo y azufre en regiones costeras (ej. Wollast et al., 1993). Por el contrario, las regiones subtropicales están claramente submuestreadas con respecto a aquellas, debido principalmente a su baja accesibilidad a la investigación oceanográfica. Existe por tanto un alto grado de incertidumbre respecto a la magnitud de los flujos de carbono en las zonas oligotróficas de los océanos. Sin embargo, su gran extensión espacial motiva que más del 50% de la productividad global del océano tenga lugar en regiones oligotróficas (Karl et al., 1996, Emerson et al., 1997); cualquier sobreestimación o subestimación en la tasas locales de producción y exportación de C tendrán por tanto una importancia crítica sobre los balances biogeoquímicos globales (Karl et al., 1996).
A diferencia de lo que ocurre en las zonas más productivas, donde la transferencia de materia está principalmente canalizada por la cadena trófica clásica, en las regiones oligotróficas tienen más importancia los procesos de reciclado dentro de las redes tróficas microbianas. Éstas se caracterizan por su elevada complejidad, sus rápidas tasas de transferencia y la coexistencia de múltiples compartimentos tróficos dentro de una misma fracción de tamaños. La dominancia de uno u otro tipo de organización trófica planctónica condiciona la velocidad de transferencia de materia y los tiempos de renovación de la masa de los distintos compartimentos, alterando de esta manera el papel que la capa superficial del océano desempeña en la circulación de materia a escala de biosfera. En este contexto, se hace necesario incrementar el nivel de comprensión acerca del efecto que la organización trófica ejerce sobre el patrón de circulación de materia y viceversa.
Organización trófica y circulación de materia
Las regiones productivas del océano presentan una distribución piramidal de la biomasa planctónica, con una amplia base de autótrofos, generalmente de tamaño grande (>20 µm), que sostiene niveles progresivamente menores de biomasa de heterótrofos. En zonas oligotróficas, por el contrario, se observan evidencias de la existencia de una pirámide invertida, en donde los componentes heterótrofos presentan una biomasa superior o equivalente a la de los autótrofos, siendo éstos generalmente de pequeño tamaño (ej. Fuhrman, 1992; Buck et al., 1996). La existencia de comunidades en las que la biomasa de heterótrofos supera a la de autótrofos ha sido explicada por la alta tasa de renovación de los autótrofos en áreas oligotróficas (Goldman et al., 1979; Baines et al., 1994). En éstas situaciones, hasta un 70 % de la producción primaria puede ser consumida por el microzooplancton (Froneman et al., 1996). Los datos existentes en el Atlántico oligotrófico muestran que el mesozooplancton puede consumir diariamente entre el 5 y el 10 % de la biomasa fitoplanctónica, mientras que consume hasta el 30 % en zonas más productivas (Huskin et al., en prep).
Los modelos conceptuales que relacionan el tipo de estructura de biomasa, el tipo de organización trófica y las vías prioritarias de transferencia de materia a través del ecosistema planctónico indican que, en sistemas caracterizados por la predominancia de redes tróficas microbianas, un porcentaje significativo de la materia circula a través del compartimento orgánico disuelto ej. (Legendre y Rassoulzadegan, 1996). Sin embargo, el nivel de conocimiento empírico que se tiene en la actualidad acerca de los fujos de materiales asociados a la fracción orgánica disuelta es escaso, y ni siquiera existen modelos conceptuales que permitan comprender de forma sintética la dinámica de las rutas ligadas a este compartimento y aun menos del posible control hidrodinámico de la misma. Algunas evidencias empíricas indican, sin embargo, que los flujos asociados con el componente disuelto deben ser importantes. Así, la magnitud de la producción de carbono y nitrógeno orgánico disuelto (COD y NOD) oscila entre un 20 y un 40 % de la producción primaria (ej. Baines y Pace, 1991; Bronk et al., 1994).
La conjunción en las regiones oligotróficas de una relativamente elevada producción de COD y una reducida mezcla vertical de la columna de agua explican la acumulación de COD que se observa en estas regiones (Karl et al., 1995), carbono que puede transportarse hacia capas subsuperficiales mediante procesos advectivos (Carlson et al., 1994). Los procesos de exportación de COD explicarían los desajustes detectados en algunas regiones del océano en los balances de carbono entre la producción primaria y la magnitud de los flujos de salida bióticos y abióticos, detectándose discrepancias de hasta un 70 % (Michaels et al., 1994). A la luz de estos resultados, resulta evidente que, dado su potencial como mecanismo de exportación de C fuera de la capa fótica, es crítico cuantificar la dinámica del COD en las zonas oligotróficas.
La acumulación de COD en las capas superiores del océano oligotrófico se derivaría, en cualquier caso, de una incapacidad de la comunidad de heterótrofos, a pesar de su elevada biomasa, para procesar instantáneamente todo el COD producido. Es decir, dicha acumulación se derivaría de un desfase en los ciclos de producción y consumo de la materia orgánica, lo cual puede relacionarse con las observaciones de pirámides invertidas, no sólo de biomasa, sino también de metabolismo neto en regiones oligotróficas (Serret et al., en prep), así como con su variabilidad. Estudios recientes indican que la eficiencia de crecimiento de las bacterias es significativamente menor en regiones oligotróficas que en zonas productivas (del Giorgio et al., 1997). Es decir, la fracción de la producción bacteriana que se canaliza hacia procesos que producen CO2 aumenta a lo largo de un gradiente autotrofía-oligotrofía, dando lugar a situaciones extremas en sistemas oligotróficos, en los que la tasa de respiración bacteriana excede a la producción primaria. Estos resultados engarzan con el reciente debate acerca del "estado trófico" del océano y más concretamente con el posible comportamiento heterótrofo del océano oligotrófico (Williams, 1998; Duarte y Agustí, 1998). En este contexto, el estudio de la dinámica del COD y, en concreto de los mecanismos que controlan el tiempo de procesado de la misma, es una prioridad para determinar las escalas espacio-temporales a las que el "estado trófico" del océano puede ser estimado a partir del balance neto del metabolismo planctónico.
Si la tasa de producción de NOD en sistemas oligotróficos fuera también elevada en términos relativos, el f-ratio calculado hasta el momento en muchos sistemas estaría subestimado. Es decir, la capacidad del ecosistema oceánico para retener (como producción exportada), a escalas de tiempo grandes, materia orgánica en forma de partículas que sedimentan (Eppley y Peterson, 1979) y materiales disueltos que difunden y son transportados hacia las capas profundas del océano (Toggweiler, 1989), sería mayor de lo que actualmente se cree. La puesta a punto de métodos que permiten la medida del enriquecimiento isotópico en 15N de la materia orgánica disuelta (Bronk y Glibert, 1991, Slawyk y Raimbault, 1995), hace posible abordar este tipo de problemas y, de esta manera, cuantificar la magnitud de la subestimación en el f-ratio.
Por otro lado, la confirmación de la existencia de situaciones heterotróficas en regiones oligotróficas obligaría a un replanteamiento del concepto de producción nueva o de las escalas significativas para su estimación. Si dicha heterotrofía resulta del alejamiento de los procesos de producción y consumo de materia orgánica, resultado y origen de la acumulación de COD, entonces los procesos de regeneración no representarían únicamente un bucle a la producción actual sino, al menos en parte, también una fuente de nutrientes nuevos a las escalas habitualmente consideradas. Por otro lado, si dicho alejamiento (en el espacio o en el tiempo) constituye una característica intrínseca del ecosistema, que estaría constituído por fases o regiones autotróficas y otras heterotróficas, la cuantificación de la máxima biomasa extraíble para mantener la integridad del sistema sólo tendría sentido a una escala que comprenda la conexión de los distintos subsistemas
La necesidad de utilizar escalas espaciales oceánicas
El cuerpo de conocimiento actual acerca de la interacción entre los modelos de organización trófica planctónica y los patrones de circulación de materia, comentado en los párrafos anteriores, está construído a partir de observaciones realizadas generalmente en escalas espaciales reducidas, en escalas temporales cortas o bien en una conjunción de ambas. Sólo podrán alcanzarse avances significativos en la comprensión del papel que la biota marina desempeña en la regulación de los ciclos biogeoquímicos globales mediante el estudio de los flujos de materiales entre compartimentos, con un enfoque integrado y en escalas espaciales relevantes en el contexto de los cambios ambientales a escala de biosfera.
Las investigaciones que adoptan este tipo de escala espacial en el campo de la oceanografía biológica han sido hasta la fecha muy escasas. Sin embargo, desde 1995, el programa "Atlantic Meridional Transect" (AMT), (http://www1.npm.ac.uk/amt/), financiado por el "National Environmental Research Council" (NERC) del Reino Unido y la NASA desarrolla anualmente campañas entre el Reino Unido y las Islas Malvinas, utilizando los tránsitos del buque oceanográfico RRS James Clark Ross. Uno de los objetivos principales del programa AMT es aprovechar la gran variabilidad geográfica de las zonas muestreadas para investigar la distribución, abundancia y actividad de los componentes del plancton y determinar el efecto de los procesos biológicos en la capa eufótica tanto sobre la producción y exportación de carbono orgánico como sobre el flujo de CO2 entre el océano y la atmósfera. El transecto AMT cruza 6 de las provincias biogeoquímicas definidas por Longhurst (1995), lo que en principio posibilita la caracterización del funcionamiento del ecosistema planctónico y la obtención de balances de materiales en regiones sometidas a diferentes regímenes de circulación y mezcla vertical.
Desde el punto de vista de la dinámica de las cadenas tróficas en diferentes regiones del Oceáno Atlántico, la participación anterior de los grupos solicitantes en el programa AMT ha permitido establecer la validez a gran escala latitudinal de algunos patrones observados previamente en escalas geográficas más limitadas:
Estos resultados han permitido por tanto la cuantificación de masas y flujos determinados que afectan a aspectos concretos del funcionamieno del ecosistema pelágico. Sin embargo, la validación y cuantificación en su conjunto del modelo conceptual derivado de estas observaciones requiere el estudio simultáneo de los flujos de carbono y nitrógeno entre los distintos compartimentos del ecosistema. Este es el único modo de comprender la relación entre la estructura trófica planctónica y el efecto neto del océano oligotrófico sobre el intercambio neto de CO2 entre el océano y la atmósfera.
Referencias
Baines, S.R., Pace, M.L. 1991. The production of dissolved organic matter by phytoplankton and its importance to bacteria: patterns across marine and freshwater ecosystems. Limnol. Oceanogr. 36: 1078-1090.
Baines, S. R., Pace, M.L., Karl, D.M. 1994. Why does the relationship between sinking flux and planktonic primary production differ between lakes and oceans? Limnol. Oceanogr.. 39: 213-226.
Bronk, D.A., Glibert, P.M. 1991. A 15N tracer method for the measurement of dissolved organic nitrogen release by phytoplankton. Mar. Ecol. Prog. Ser. 77: 171-182.
Bronk, D.A., Glibert, P.M., Ward, B.B. 1994. Nitrogen uptake, dissolved organic nitrogen release, and new production. Science. 265: 1843-1846.
Buck, K. R., Chavez, F.P., Campbell, L. 1996. Basin-wide distributions of living carbon components and the inverted trophic pyramid of the central gyre of the North Atlantic Ocean, summer 1993. Aquatic Microb. Ecol. 10: 283-298.
Carlson, C.A., Ducklow, H.W., Michaels, A.F. 1994. Annual flux of dissolved organic carbon from the euphotic zone in the northwestern Sargasso Sea. Nature. 371: 405-408.
del Giorgio, P.A., Cole, J.J., Cimbleris, A. 1997. Respiration rates in bacteria exceed phytoplankton production in unproductive aquatic systems. Nature 385:148-151.
Duarte, C., Agustí, S. 1998. The CO2 balance of unproductive aquatic ecosystems. Science. 281: 234-236.
Emerson, S., P. Quay, D. Karl, C. Winn, L. Tupas, M. Landry. 1997. Experimental determination of the organic carbon flux from open-ocean surface waters. Nature. 389: 951-954.
Eppley, R.W., Peterson, B.J. 1979. Particulate organic matter flux and planktonic new production in the deep ocean. Nature, 282: 677-680.
Falkowski, P. G., Barber, R. T., Smetacek, V. 1998. Biogeochemical controls and feedbacks on ocean primary production. Science, 281, 200-206.
Field, C. B., Behrenfeld, M. J., Randerson, J. T., Falkowski, P. 1998. Primary production of the biosphere: integrating terrestrial and oceanic components. Science, 281, 237-240.
Fernández, E., Teira, M.J. Pazó, N. González, P. Serret. (en preparación). Release of dissolved organic carbon by microplankton in the Atlantic Ocean: relation to primary and net community production.
Fuhrman, J. 1992. Bacterioplankton roles in cycling of organic matter: The microbial food web. En: Primary Productivity and Biogeochemical Cycles in the Sea. Ed. P.G. Falkowski y A.D. Woodhead. Plenum Press. 361-383.
Goldman, J. C., McCarthy, J.J., Peavy, D.G. 1979. Growth rate influence on the chemical composition of phytoplankton in oceanic waters. Nature. 279: 210-215.
Holligan, P. M. 1992. Do marine phytoplankton influence global climate? En P. G. Falkowski y A. D. Woodhead (Eds), Primary productivity and biogeochemical cycles in the sea. Plenum Press, New York.
Karl, D.M., Letelier, R.M., Hebel, D., Tupas, L., Dore, J., Christian, J., Winn, C. 1995. Ecosystem chages in the North Pacific subtropical gyre attributed to the 1991-1992 El Niño. Nature 373:230-234.
Karl, D. M., Christian, J. R., Dore, J. E., Hebel, D. V., Letelier, R. M., Tupas, L. M. Winn, C. W. 1996. Seasonal and interannual variability in primary production and particulate flux at Station ALOHA. Deep-Sea Res., 43, 539-568.
Legendre, L., Rassoulzadegan, F. 1996. Food-web mediated export of biogenic carbon in oceans. Mar.Ecol.Prog.Ser. 145: 179-193.
Longhurst, A. Sathyendranath, S., Platt, T., Caverhill, C. 1995. An estimate of global primary production in the ocean from satellite radiometer data. J.Plankton Res. 17(6): 1245-1271.
Marañón, E., Holligan, P. M., Varela, M., Mouriño, B., Bale, A.J. 1999. Basin scale variability of phytoplankton biomass, production and growth in the Atlantic Ocean. Deep-Sea Res. (en prensa).
Marañón, E. y otros. (en preparación). Temporal variability in the latitudinal distribution of size-fractionated chlorophyll and primary production in the Atlantic Ocean.
Martin, J. H. y otros. 1994. Testing the iron hypothesis in ecosystems of the equatorial Pacific Ocean. Nature, 371, 123-129.
Michaels, A.F., Bates, N.R., Buesseler, K.O., Carlson, C.A., Knap, A.H. 1994. Carbon-cycle imbalances in the Sargasso Sea. Nature. 372: 537-540.
Sarmiento, J. L., Bender, M. 1994. Carbon biogeochemistry and climate change. Photosynthesis research, 39: 209-234.
Serret, P., Robinson, C., Tilstone, G., Teira, E., Fernández, E. (En preparación). Basin scale distribution of microplankton production and respiration balances in the eastern Atlantic Ocean (35 °S-50 °N).
Slawyk, G., Raimbault, P. 1995. Simple procedure for simultaneous recovery of dissolved inorganic and organic nitrogen in 15N-tracer experiments and improving the isotopic mass balance. Mar. Ecol. Prog. Ser., 124: 289-299.
Toggweiler, J.R. 1989. Is the downward dissolved organic matter (DOM) flux important in carbon transport ? En: Berger, W.H., Smetacek, V.S. y Wefer, G. (eds), Productivity in the ocean: Past and present. Wiley, pp. 65-83.
Volk, T., Hoffert, M. I. 1985. Ocean carbon pumps: analysis of relative strengths and efficiencies in ocean-driven atmospheric CO2 changes. In: The carbon cycle and atmospheric CO2: natural variations archean to present, E. Sundquist and W. S. Broecker (Eds.), Geographical Monograph 32, American Geophysical Union.
Williams, P.J. leB. 1998. The balance of plankton respiration and photosynthesis in the open oceans. Nature. 394: 55-57.
Williamson, P., Holligan, P. M. 1990. Ocean productivity and climate change. Trends Ecol. Evol., 5(9): 299-303.
Wollast, R., McKenzie F.T., Chou, L. 1993. Interactions of C, N, P and S biogeochemical cycles and global change. NATO ASI Series. Serie I. Global Environmental Change. Springer-Verlag. Berlin. 521 pp.
Woods, J, Barkmann, W. 1993. The plankton multiplier—positive feedback in the greenhouse. J. Plankton Res., 15: 1053-1074.
OBJETIVOS
Descripción realista de los objetivos concretos del proyecto. Se valorará la adecuación de la propuesta a las prioridades del correspondiente Programa Nacional, así como la relevancia de los objetivos en el contexto científico-técnico correspondiente, de acuerdo con la modalidad a la que se presenta la solicitud.
Recuerde que la Modalidad A se refiere a proyectos para la generación de conocimientos científicos y tecnológicos que contribuyan al desarrollo de los sectores productivos o a la mejora de la planificación de las diferentes áreas socioeconómicas. La Modalidad B se refiere a proyectos orientados a la aplicación tecnológica a corto plazo, para mejorar la eficacia y competitividad de los sectores industriales, de servicios o unidades de las administraciones públicas, o para la mejor gestión de los recursos naturales.
El proyecto CIRCANA tiene como meta:
Avanzar en la comprensión del efecto que el componente biótico
del océano ejerce sobre procesos biogeoquímicos relevantes
en el contexto del cambio global
Se pretende integrar en un marco conceptual general una serie de resultados, ya comentados en el apartado anterior, que se han venido obteniendo durante las últimas décadas mediante observaciones del océano abierto realizadas en escalas geográficas reducidas:
La integración de estas observaciones en un modelo conceptual general permite generar escenarios hipotéticos dispuestos a lo largo de un continuo de situaciones que discurren desde sistemas claramente oligotróficos hasta aquellos caracterizados por una elevada productividad.
Modelo conceptual hipotético
En sistemas oligotróficos como los giros subtropicales, la relación biomasa heterótrofa/biomasa autótrofa es más elevada que en sistemas productivos. Esta estructura se sustenta en la elevada tasa de renovación de la biomasa del picoplancton. El tipo de organización trófica resultante da lugar a un elevado flujo de materia fotoasimilada hacia el compartimento disuelto, lo cual tiene tres consecuencias:
La importancia cuantitativa del picoplancton motiva que la relación entre la biomasa de microzooplancton y la de mesozooplancton sea elevada. En esta situación la contribución potencial del mesozooplancton al metabolismo planctónico sería muy reducida. A su vez, la desviación de un porcentaje elevado de la producción primaria hacia la fracción disuelta reduce la cantidad de materia disponible para el microzooplancton. Las interrelación de los procesos anteriores da lugar a balances producción/respiración generalmente menores en sistemas oligotróficos con respecto a aquellos de elevada productividad.
Objetivo general
Las actividades planteadas en este proyecto pretenden evaluar el grado de veracidad del modelo conceptual planteado. Con este fin se define el objetivo general del proyecto, que es:
Determinar la magnitud de los flujos de carbono y nitrógeno asociados a los tipos de organización trófica planctónica característicos de las diferentes regiones del Océano Atlántico
Objetivos particulares
Este objetivo general se desglosa en una serie de objetivos particulares relacionados con el modelo conceptual planteado. Para cada una de las regiones del océano Atlántico se pretende cuantificar:
Referencias
Baines, S. R., Pace M.L., Karl, D.M. 1994. Why does the relationship between sinking flux and planktonic primary production differ between lakes and oceans? Limnol. Oceanogr.. 39: 213-226.
Buck, K.R., Chavez, F.P., Campbell, L. 1996. Basin-wide distributions of living carbon components and the inverted trophic pyramid of the central gyre of the North Atlantic Ocean, summer 1993. Aquat.Microb.Ecol. 10:283-298.
del Giorgio, P.A., Cole, J.J., Cimbleris, A. 1997. Respiration rates in bacteria exceed phytoplankton production in unproductive aquatic systems. Nature 385:148-151.
Duarte, C., Agustí, S. 1998. The CO2 balance of unproductive aquatic ecosystems. Science. 281: 234-236.
Fernández, E., Teira, M.J. Pazó, N. González, P. Serret. (en preparación). Release of dissolved organic carbon by microplankton in the Atlantic Ocean: relation to primary and net community production.
Froneman, P.W., Perissinotto, R., McQuaid, C.D. 1996. Seasonal variations in microzooplankton grazing in the region of the subtropical convergence. Mar. Biol. 126: 433-442.
Fuhrman, J. 1992. Bacterioplankton roles in cycling of organic matter: The microbial food web. En: Primary Productivity and Biogeochemical Cycles in the Sea. Ed. P.G. Falkowski y A.D. Woodhead. Plenum Press. 361-383.
Huskin, I., Anadón, R., Wood-Walker, R, Head, B., Haris, R.M., Robins, D. (en preparación). Grazing impact of mesozooplankton in the Atlantic at a biogeochemical provinces scale.
Karl, D.M., Letelier, R.M., Hebel, D., Tupas, L., Dore, J., Christian, J., Winn, C. 1995. Ecosystem chages in the North Pacific subtropical gyre attributed to the 1991-1992 El Niño. Nature 373:230-234.
Legendre, L., Rassoulzadegan, F. 1996. Food-web mediated export of biogenic carbon in oceans. Mar.Ecol.Prog.Ser. 145: 179-193.
BENEFICIOS DEL PROYECTO
En las propuestas de la Modalidad A:
ë Utilidad de la propuesta para los sectores socioeconómicos a los que se dirige. Explicar el beneficio según su cobertura (nacional, local, sectorial).
ë En su caso, documentación que justifique la colaboración de uno o varios EPO (empresa, entidad social o unidad de la administración que declara su interés por el proyecto y, en consecuencia, se compromete a colaborar o hacer un seguimiento de su progreso). El papel asignado al EPO conlleva como mínimo su compromiso con el seguimiento y valoración de los beneficios esperados en la ejecución del proyecto. La Dirección General de Enseñanza Superior e Investigación Científica podrá solicitar, directamente al representante del EPO, informe confidencial sobre estos aspectos.
En las propuestas de la Modalidad B:
ë Previsible obtención de resultados con posibilidad de transferencia a sectores empresariales o sociales, con especial referencia a las pequeñas y medianas empresas y a los centros de innovación y tecnología.
El proyecto CIRCANA supone una contribución relevante al conocimiento de los patrones generales de circulación de materia en ecosistemas marinos en escalas espaciales significativas en el contexto de los cambios ambientales que acontecen en la biosfera. La dimensión del proyecto es, en consecuencia, global. Desde la perspectiva nacional, su interés proviene tanto de su vinculación con programas internacionales dentro del ámbito IGBP (JGOFS, GLOBEC, SOLAS) como de su adecuación a las acciones claves del próximo V Programa Marco de la Unión Europea.
En este sentido, las metas de CIRCANA se encuadran dentro de los objetivos generales JGOFS, "Determinar y comprender a escala global los procesos que controlan la variabilidad en los flujos de carbono y elementos asociados en relación a su intercambio con la atmósfera, el sedimento y los márgenes continentales", con los de la actividad 2.2 de GLOBEC "Determinar el papel de las interacciones entre procesos físicos y biológicos a múltiples escalas sobre la estructura de los ecosistemas marinos y en la forma en que ésta responde a cambios ambientales globales", y con los del futuro programa SOLAS, "Estudiar las interacciones entre el sistema biogeoquímico marino, la atmósfera y el clima y como este sistema afecta y es afectado por cambios ambientales y climáticos pasados y futuros".
La información derivada de CIRCANA es, además, de especial relevancia en el marco de la acción clave 2.2.2 del futuro V Programa Marco de la Unión Europea, cuyo objetivo es: "Aumentar el conocimiento acerca de las interacciones existentes entre ecosistemas y los ciclos de carbono y nitrógeno, incluyendo específicamente los ecosistemas marinos".
Desde la perspectiva científica, la innovación que supone la "aproximación AMT" es similar a la que con anterioridad se ha adoptado en el campo de la oceanografía química (GEOSECS) o aún en la actualidad en el marco de la oceanografía física (WOCE) y estriba principalmente en la gran cobertura espacial que alcanza y en la posibilidad de obtener información en dos periodos contrastados del ciclo estacional: primavera y otoño.
Desde el punto de vista logístico, la realización del proyecto dentro del ámbito del proyecto AMT, utilizando el buque RRS James Clark Ross como plataforma, representa un valor añadido sustancial al permitir realizar con un bajo coste campañas oceanográficas en mar abierto, cubriendo grandes escalas de variabilidad espacial y aprovechando una instrumentación de primera línea para el estudio multidisciplinar del océano. Además de la disponibilidad de barco, el proyecto se beneficia directamente de toda la información generada directamente por el componente británico del programa.
CIRCANA permitirá continuar con la actividad investigadora iniciada tras la financiación por el plan nacional CYTMAR de dos acciones especiales (MAR95-1970-E y MAR98-1417-E) y, de esta manera, situar a los grupos solicitantes en una posición ventajosa de cara a las próximas convocatorias de proyectos incluídas en el V programa marco de la Unión Europea.
METODOLOGÍA Y PLAN DE TRABAJO
Concreción de los objetivos especificando metodología, actividades programadas y resultados previsibles. Se valorará el rigor en el planteamiento y la adecuada planificación temporal de las actividades.
ë El plan de trabajo ha de poner de relieve la necesidad de la ayuda que se solicita en cada concepto para alcanzar los objetivos propuestos, con especial atención al capítulo de gastos de personal y de material inventariable cuyas necesidades deben justificarse adecuadamente.
Indicar, en diagramas similares a los que se incluyen como ejemplo, el desarrollo temporal de las actividades programadas, la imputación de los posibles gastos de personal a las diferentes actividades y la distribución, por actividades, del presupuesto solicitado.
Campañas AMT: estrategia y limitaciones
El proyecto que se plantea se llevará a cabo usando como plataforma de muestreo el programa británico "Atlantic Meridional Transect" (AMT). El programa AMT se inició en septiembre de 1995 como resultado de un acuerdo entre el BAS (British Antarctic Survey) y el PML (Plymouth Marine Laboratory) para utilizar el buque oceanográfico RRS James Clark Ross durante los dos tránsitos que éste realiza anualmente entre el Reino Unido y las Islas Malvinas, siguiendo el recorrido superpuesto a la imagen SeaWifs que se adjunta. El plan de trabajo se basa en el muestreo de una estación diaria hasta 200 m de profundidad, totalizando un total de 30 estaciones por campaña a lo largo de un transecto entre 50° N y 50° S.
La gran cobertura espacial que se alcanza supone a su vez la mayor limitación del proyecto: la escasa resolución espacial y temporal del mismo. Si bien la estrategia de muestreo garantiza la construcción de los balances de biomasa y de flujo de C y N correspondientes a las distintas regiones del Atlántico, ha de tenerse en cuenta que estos balances deben interpretarse como imágenes dentro de un patrón de variación temporal continuo. Por ello, preguntas como ¿es el océano Atlántico heterotrófico? no podrán ser contestadas mientras no se disponga de información en escalas temporales adecuadas.
El acceso de los investigadores que firman esta propuesta de financiación al RRS James Clark Ross está garantizado por la dirección del proyecto AMT. En este sentido, se adjunta una carta en la cual el director del proyecto AMT, Dr. James Aiken, se compromete a facilitar los camarotes y espacio necesarios para la consecución de los objetivos planteados en CIRCANA.
Además, el componente británico del proyecto suministrará la siguiente información complementaria de relevancia para CIRCANA:
Fases del proyecto
El proyecto consta de 3 fases:
1.- Fase preparatoria y de puesta a punto de metodología
En esta primera fase se pondrán a punto los métodos de medida de aquellas variables que no hayan sido estudiadas en anteriores campañas AMT por nuestros grupos de investigación. En concreto: producción bacteriana, balances de O2 tanto del microplancton como del mesozooplancton y estimación de flujos relacionados con la circulación del nitrógeno. Tras un periodo de experimentación en el laboratorio, la fase de puesta a punto metodológica concluirá con la participación en una campaña en el otoño del 2000. Esta campaña se destinará prioritariamente a la experimentación metodológica con el fin de garantizar la adecuación de las metodologías a las características de las diferentes regiones estudiadas.
La elección de esta fecha para la primera campaña permite reducir al máximo los costes de transporte de material y equipos ya que en septiembre el transecto se inicia en Gran Bretaña.
A lo largo de esta fase se construirá la página Web CIRCANA en la que se ubicará toda la información referente al proyecto así como la base de datos del mismo.
2.- Fase de construcción de los balances de biomasa y flujos de C y N
La cuantificación simultánea de todas las masas y flujos de carbono y nitrógeno necesarios para la consecución de los objetivos del proyecto se llevará a cabo en dos campañas a celebrar en:
La realización de dos campañas que comparten el mismo planteamiento se fundamenta en el interés de obtener información, aunque limitada, sobre la variabilidad temporal de las biomasas y flujos estudiados.
3.- Fase de elaboración de resultados y redacción de trabajos científicos
Tras la realización de las campañas, el esfuerzo se concentrará en tres aspectos:
Esta fase concluirá con la organización de una reunión general en la que participarán todos los investigadores del proyecto. El objetivo de esta reunión general, que tendrá estructura de "workshop", es la puesta en común de los resultados y la discusión acerca de la elaboración de un trabajo de síntesis.
La ubicación temporal de las fases del proyecto se detalla en el diagrama de tiempos que se presenta a continuación.
|
PLAN DE ACTIVIDADES · DIAGRAMA DE TIEMPOS |
|||||||||||||||||||||||||||||||||||||||
|
Actividad |
AÑO 1 |
AÑO 2 |
AÑO 3 |
||||||||||||||||||||||||||||||||||||
|
E |
F |
M |
A |
M |
J |
J |
A |
S |
O |
N |
D |
E |
F |
M |
A |
M |
J |
J |
A |
S |
O |
N |
D |
E |
F |
M |
A |
M |
J |
J |
A |
S |
O |
N |
D |
||||
|
1ª Reunión de coordinación |
|||||||||||||||||||||||||||||||||||||||
|
Puesta a punto de metodología |
|||||||||||||||||||||||||||||||||||||||
|
Construcción de la página web |
|||||||||||||||||||||||||||||||||||||||
|
1ª Campaña |
|||||||||||||||||||||||||||||||||||||||
|
2ª Reunión de coordinación |
|||||||||||||||||||||||||||||||||||||||
|
2ª Campaña |
|||||||||||||||||||||||||||||||||||||||
|
3ª Campaña |
|||||||||||||||||||||||||||||||||||||||
|
Análisis de muestras |
|||||||||||||||||||||||||||||||||||||||
|
Construcción de la base de datos |
|||||||||||||||||||||||||||||||||||||||
|
Elaboración de resultados |
|||||||||||||||||||||||||||||||||||||||
|
Redacción de trabajos científicos |
|||||||||||||||||||||||||||||||||||||||
|
Workshop de síntesis |
|||||||||||||||||||||||||||||||||||||||
|
Redacción del informe final |
|||||||||||||||||||||||||||||||||||||||
Relación de actividades
La consecución de los objetivos descritos en el apartado anterior requiere la medida simultánea de numerosas variables que en conjunto definen el balance de biomasas y flujos del sistema planctónico tal como se muestra en el esquema adjunto. Los códigos indican cada una de las variables que serán medidas, así como el subproyecto encargado de su cuantificación.
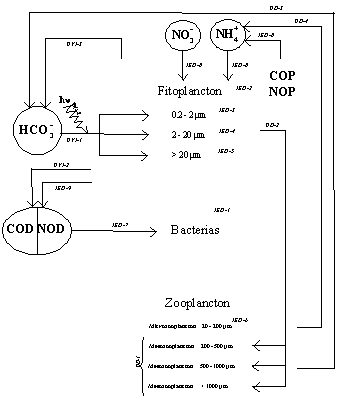
Esquema de las variables de biomasa y de flujo que serán cuantificadas en CIRCANA. Los códigos de las variables indican el subproyecto responsable de su cuantificación, tal como se detalla en la tabla de la página siguiente. UVI - Universidad de Vigo (Subproyecto 1), IEO - Instituto Español de Oceanografía (Subproyecto 2), UO - Universidad de Oviedo (Subproyecto 3).
Metodología
UVI-1 : Producción primaria fraccionada por tamaños
Investigador responsable: Emilio Marañón
En cada estación se tomarán por triplicado muestras de agua de 6 profundidades y se inocularán con 20 m Ci de NaH14CO3. Las muestras se mantendrán en incubadores de cubierta que simulan la intensidad y espectro de radiación correspondiente a las profundidades de origen de las muestras. La temperatura de incubación se mantendrá también similar a la de origen de las muestras mediante el empleo de refrigeradores de elevada capacidad. Las muestras se filtrarán a través de filtros de policarbonato de 0.2, 2 y 20 m m de diámetro de poro. El procedimiento a seguir con los filtros será el descrito en Marañón et al. (1999) y en los protocolos JGOFS (UNESCO, 1994).
UVI-2 : Producción de carbono orgánico disuelto
Investigador responsable: Emilio Marañón
En todas las estaciones, y en cuatro profundidades, se tomarán por cuadriplicado muestras de 35 ml, para la determinación de la tasa de producción de carbono orgánico disuelto por el microplancton siguiendo básicamente el método descrito por Mague et al. (1980). Las muestras se inoculan con 35-50 m Ci de NaH14CO3, dependiendo de la biomasa fitoplanctónica de la muestra. Tras 2 horas de incubación en incubadores de cubierta como los mencionados en el apartado anterior, las muestras se filtran a través de filtros de policarbonato de 0.2 m m de diámetro de poro utilizando un equipo especialmente diseñado para la recolección del filtrado. Los filtros se procesan tal como se describe en el epígrafe anterior. El filtrado se acidifica con 40 µl de HCl al 50 % y se burbujea durante 24 horas con aire que previamente atraviesa una trampa de atmósfera alcalina con el objeto de eliminar cualquier contaminación por CO2 marcado con 14C. Tras la fase de descontaminación el filtrado se procesa de forma similar a los filtros.
UVI-3 : Producción y consumo de oxígeno por el microplancton
Investigador responsable: Emilio Fernández
En todas las estaciones y de 6 profundidades se tomarán muestras de agua para la determinación de las tasas de producción y consumo de oxígeno. De cada profundidad se llenarán 12 muestras de 125 ml en botellas de borosilicato individualmente calibradas. Cuatro de las botellas se fijan inmediatamente. Las ocho restantes se mantienen durante 24 h en un incubador de características similares a las descritas en el apartado de producción primaria, cuatro de ellas en oscuridad y las cuatro restantes bajo un ciclo luz-oscuridad. La concentración de oxígeno disuelto se medirá mediante el método de Winkler con un titulador automático Metrohm DMS Titrino, adoptando un punto final potenciométrico.
UVI-4 : Relaciones fotosíntesis-irradiancia y absorción de luz
Investigador responsable: Ramiro Varela
En todas las estaciones y en tres profundidades, se realizarán experimentos para determinar los parámetros de las relaciones fotosíntesis-irradiancia de las poblaciones de fitoplancton. De cada una de las profundidades se llenarán 14 botellas de 80 ml de capacidad, que se inocularán con 10 µCi de NaH14CO3. Éstas se colocarán en incubadores que generan un gradiente de irradiancia entre 1500 y 8 µE m-2 s-1. Tras dos horas de incubación, las muestras se filtran a través de filtros de fibra de vidrio que se procesan tal como se detalla en epígrafes anteriores. A partir de las relaciones fotosíntesis-irradiancia se calcularán los parámetros fotosintéticos Pmax, a y b (en el caso de detectarse fotoinhibición).
En las mismas estaciones y profundidades, se realizarán mediciones de la absorción de luz por el material particulado presente en la columna de agua. Para cada profundidad se filtrará entre 1 y 3 litros de agua de mar por filtros de 0.2, 2 y 20 mm de diámetro de poro. La densidad óptica de los filtros se medirá en un espectrofotómetro y se convertirá en absorción utilizando el método de Bricaud y Stramski (1990). El material retenido en los filtros se sumergirá en una solución de metanol al 100% durante 3 períodos de 30, 10 y 10 minutos respectivamente para la eliminación de los pigmentos presentes. Una nueva lectura en el espectrofotómetro proporcionará la absorción debida al material no pigmentado (Kishino et al., 1985) obteniéndose por diferencia la correspondiente al material algal. Este método empírico será desarrollado en paralelo al procedimiento matemático de separación en las fracciones correspondientes al material algal y no algal a partir de la absorción total (Varela et al., 1998)
IEO-1 : Biomasa bacteriana
Investigador responsable: Ángeles Cid
Se determinará la abundancia de bacterias mediante citometría de flujo. Las muestras recogidas en cada una de las estaciones de muestreo se fijarán con una mezcla de paraformaldehido y glutaraldehido, para ser posteriormente congeladas en nitrógeno líquido. Las muestras se mantendrán congeladas hasta su análisis en el laboratorio (al final de la campaña). Para el recuento de bacterias se utilizará el protocolo de tinción con SYTO 13 de del Giorgio et al. (1996). En algunas muestras se hará una comparación de los contajes realizados mediante microscopía de epifluorescencia en alícuotas filtradas sobre filtros de policarbonato (0.2 µm de tamaño de poro) y teñidas con DAPI (Porter y Feig, 1980).
IEO-2 : Clorofilas a,b y c fraccionadas por tamaños
Investigador responsable: Manuel Varela
En cada estación se tomarán muestras de agua de 250 ml de 7 profundidades que se filtrarán por filtros de policarbonato de 0.2, 2 y 20 m m. Los pigmentos liposolubles se extraerán en acetona 90 % y la concentración de las clorofilas a, b y c se cuantificará espectrofluorimétricamente con un equipo Safas flx. El almacenamiento, extracción, lectura fluorométrica y cálculo de resultados se efectuará siguiendo las directrices expuestas en los protocolos JGOFS (UNESCO, 1994).
IEO-3 : Biomasa y composición de picofitoplancton
Investigador responsable: Ángeles Cid
Se determinará la abundancia de flagelados fotosintéticos, proclorófitos y cianobacterias mediante citometría de flujo. Las muestras se recogerán, conservarán y procesarán como se describe en el caso de la biomasa bacteriana.
IEO-4 : Biomasa y composición de nanofitoplancton
Investigador responsable: Manuel Varela
Los organismos fitoplanctónicos menores de 20 µm se muestrearán con botellas Niskin a diferentes profundidades de la zona fótica. En cada profundidad se tomarán dos muestras: una de ellas se fijará con formol neutralizado, para el estudio de los cocolitoforidos, y otra con solución de Lugol, para el resto de organismos. La identificación y recuento de los organismos se llevará a cabo siguiendo la técnica de Utermöhl (1957). Para cada especie se calculará su volumen celular, a partir del cual se estimará su contendio en carbono, utilizando para ello las ecuaciones descritas en la bibliografía (Strathman, 1967; Eppley et al., 1970; Smetacek, 1975).
IEO-5 : Biomasa y composición de microfitoplancton
Investigador responsable: Santiago Fraga
Los organismos de fitoplancton de mayor tamaño (20-200µm) se recogerán mediante pescas verticales (0-200 m) con malla de 20 µm. Para cuantificar la biomasa de este componente se procederá de la misma manera que con los organismos pequeños. A partir de los volúmenes celulares se estimarán los diametros esféricos medios (DEM, Malone, 1980), y la distribución de las biomasas por clases de tamaño. Se prestará especial atención a la cuantificación de la biomasa de los organismos fijadores de nitrógeno.
IEO-6 : Biomasa y composición de microzooplancton
Investigador responsable: Ana Miranda
El análisis de la biomasa y composición del microzooplancton (fracción entre 20 y 200 µm) se realizará sobre las mismas muestras que el microfitoplancton. La estimacion de biomasa se hará por análisis elemental CNH, filtrando a bordo del barco y congelando el filtro. Se calcularán posteriormente biovolúmenes de microzooplancton con el fin de estimar biomasa heterótrofa de la fracción planctónica recogida entre 20 y 200 µm. La composición y abundancia de microzooplancton se estudiará analizando con una lupa binocular las muestras fijadas con formol neutralizado al 5% en agua de mar, tras filtración por una malla de 200 µm.
IEO-7 : Producción bacteriana
Investigador responsable: Ángeles Cid
La producción heterotrófica bacteriana se determinará mediante la técnica de incorporación de leucina marcada con 3H (Kirchman et al., 1985), modificada como micrométodo por Smith & Azam (1992).
IEO-8 y 9 : Consumo y regeneración de NID y producción de NOD
Investigador responsable: Antonio Bode
Se realizarán incubaciones de agua oceánica de tres profundidades por estación (superficie y las profundidades ópticas correspondientes al 10% y 1% de la irradiancia incidente en superficie) en botellas de policarbonato a las que se añadirá nitrato y amonio marcados con 15N para determinar las tasas brutas de incorporación de nitrógeno inorgánico disuelto (NID) y la tasa de liberación de nitrógeno orgánico disuelto (NOD). El procedimiento analítico a seguir será el propuesto por Slawyk et al., (1998), que implica el análisis del enriquecimiento isotópico del NID y del nitrógeno orgánico total (NOT = NOD + NOP). Las incubaciones se realizarán a bordo utilizando un sistema refrigerado con agua de superficie, que simula los niveles de irradiancia existentes en el lugar de muestreo. Al terminar las incubaciones se recogerán dos alícuotas de cada botella, una de ellas sin filtrar para el análisis del NOT y otra filtrada para el análisis del NOD, que se conservarán fijadas con HgCl2 hasta su procesamiento en el laboratorio. Las determinaciones en el laboratorio se basarán en la conversión de todo el nitrógeno de las botellas (NID, NOD y NOT) a nitrato (mediante oxidación con K2S2O8) y finalmente a amonio (mediante alcalinización con MgO), y su recogida mediante difusión en tiras de fibra de vidrio para su secado y análisis del enriquecimiento relativo en 15N (mediante un espectrómetro de masas). La medida de las concentraciones de nutrientes, NOT y NOD se realizará mediante el análisis de amonio y nitrato en un autoanalizador (Grasshoff et al., 1983).
UO-1 : Biomasa y composición de mesozooplancton
Investigador responsable: Florentina Álvarez
La biomasa y composición de mesozooplancton se obtendrá a partir de muestras integradas entre 0 y 200 m, recogidas mediante una red doble WP2, siguiendo la metodología usada en campañas AMT precedentes. La muestra de un colector se dividirá en dos fracciones iguales mediante un separador Folsom. La muestra para estima de biomasa se fraccionará en tres clases de tamaño mediante el uso de tres mallas de 200, 500 y 1000 µm de tamaño. Se obtendrá el peso seco y el contenido en carbono y nitrógeno del zooplancton mediante pesada el primero, y mediante un Autoanalizador Elemental CNH los segundos. La muestra destinada al estudio de la composición específica y estima de abundancias se fijará mediante formol al 4 % tamponado con bórax. En el laboratorio se fraccionará en las tres clases mencionadas en el párrafo anterior, identificándose los organismos hasta el nivel de especie. La misma muestra utilizada para la determinación de los organismos se utilizará para determinar la estructura de tamaños del mesozooplancton, para lo que se filmaran varios campos de la muestra mediante cámara de video SONY SSC-C370P. Las imágenes se digitalizaran y se medirán la longitud y anchura de los organismos, medidas con las que se calculará el VEE (volumen esférico equivalente) y posteriormente el espectro de tallas
UO-2 : Tasas de ingestión del zooplancton
Investigador responsable: José Luis Acuña
Las tasas de ingestión del microzooplancton se estimarán a partir de los espectros de talla del microzooplancton obtenidas en el apartado IEO6, utilizando las relaciones alométricas definidas para el zooplankton por Hansen et al (1997). Las tasas de ingestión del mesozoplancton se obtendrán por el método de la fluorescencia del contenido digestivo Mackas y Bohrer (1976), ya previamente utilizado en las campañas AMT por Huskin et al., (en prep.). Los contenidos se estudiarán en las tres clases de tamaño ya mencionadas. Las tasas de evacuación han sido obtenidas previamente para los copépodos por Huskin et al (op. cit.), por lo que sólo se realizaran algunos experimentos complementarios. En el caso de las Apendicularias se usará el tiempo de paso estimado por López-Urrutia y Acuña (sometido), y en el de las salpas los valores indicados por Madin y Cetta (1984). Los valores de ingestion total del zooplancton se obtendrán multiplicando la ingestion individual por la abundancia de individuos de cada clase obtenidos, y se compararan con las estimadas a partir de las ecuaciones alométricas de Hansen et al (op. cit.)
UO-3 : Consumo de oxígeno por el mesozooplancton
Investigador responsable: Ricardo Anadón
Las tasas de consumo de oxígeno del microzooplancton se estimaran a partir de la abundancia y talla de los organismos determinada en el apartado IEO6, utilizando la ecuación alométrica de Ikeda (1985) para la respiración en función de la temperatura. La tasa respiratoria del mesozooplancton se obtendrá mediante incubaciones en agua filtrada de individuos de la totalidad de la comunidad recogidos con una red WP2, y fraccionados en tres clases de tamaño mediante filtración inversa. Se realizarán cuatro réplicas por tamaño, y el tiempo de incubación será de 24 horas. La temperatura de incubación corresponderá a la del máximo de clorofila. El oxígeno disuelto en el agua se analizará por el método de Winkler (Grasshoff et al 1983) al principio y al final de la incubación, mediante un titulador automático Metrohm 716 DMS Titrino. De los organismos incubados se obtendrá el contenido en carbono y nitrógeno, y se calculará la tasa específica de respiración.
UO-4: Regeneración de amonio por el mesozooplancton
Investigador responsable: Ricardo Anadón
La medida de la tasa de regeneración de amonio por el mesozooplancton se estimará a partir del incremento en concentración de amonio en las mismas incubaciones que en el caso de la medida de la tasa de respiración. Se analizarán amonio, nitrato y fosfato antes y después de la incubación mediante los métodos descritos en Grasshoff et al. (1983).
Gestión de la base de datos
Investigador responsable: Emilio Fernández
La elaboración de la base de datos será responsabilidad de los miembros del comité de coordinación del proyecto, que se ocuparán de centralizar la información y facilitar el intercambio de datos entre diferentes grupos participantes con el objetivo de optimizar el proceso de elaboración de resultados. Con este fin, se construirá una página Web en la que se ubicará toda la información que sea de relevancia para los miembros del proyecto y servirá de ventana hacia el resto de la comunidad científica. La página Web dispondrá de una sección de acceso restringido que constituirá la base de datos del proyecto. La gestión de la página web será responsabilidad del coordinador del proyecto. En un nivel superior, esta base de datos podrá ser añadida al banco de datos de AMT, que en la actualidad es gestionado por el PML con el objetivo final de incluirlo dentro del BODC (British Oceanographic Data Centre). El mantenimiento de estas bases de datos asegura que la información sea puesta a disposición de la comunidad internacional y aumenta los beneficios científicos a medio plazo.
En la tabla siguiente se presenta un cuadro resumen en el que se detallan las variables consideradas junto a los investigadores responsables de su medida.
|
Código |
Variable |
Investigador responsable |
|
UVI-1 |
Producción en clases de tamaño |
Emilio Marañón |
|
UVI-2 |
Producción de COD |
Emilio Marañón |
|
UVI-3 |
Producción y consumo de O2 |
Emilio Fernández |
|
UVI-4 |
Relaciones fotosíntesis-irradiancia y absorción de luz |
Ramiro Varela |
|
IEO-1 |
Biomasa bacteriana |
Ángeles Cid |
|
IEO-2 |
Clorofila en clases de tamaño |
Manuel Varela |
|
IEO-3 |
Biomasa y composición de picofitoplancton |
Ángeles Cid |
|
IEO-4 |
Biomasa y composición de nanofitoplancton |
Manuel Varela |
|
IEO-5 |
Biomasa y composición de microfitoplancton |
Santiago Fraga |
|
IEO-6 |
Biomasa y composición de microzooplancton |
Ana Miranda |
|
IEO-7 |
Producción bacteriana |
Ángeles Cid |
|
IEO-8 |
Consumo y regeneración de NID |
Antonio Bode |
|
IEO-9 |
Producción de NOD |
Antonio Bode |
|
UO-1 |
Biomasa de mesozooplancton |
Florentina Álvarez |
|
UO-2 |
Tasas de ingestión por el mesozooplancton |
José Luis Acuña |
|
UO-3 |
Consumo de O2 por el mesozooplancton |
Ricardo Anadón |
|
UO-4 |
Regeneración de NH4 por el mesozooplancton |
Ricardo Anadón |
Referencias
Bricaud, A., Stramski, D. 1990. Spectral absorption coefficients of living phytoplankton and non algal biogenous matter: A comparison between the Peru upwelling area and the Sargasso Sea. Limnol. Oceanogr. 35: 562-582.
Del Giorgio, P.A., Bird, D.F., Prairie, Y.T., Planas, D. 1996. Flow cytometric determination of bacterial abundance in lake plankton with the green nucleic acid stain SYTO 13. Limnol. Oceanogr. 41: 783-789.
Eppley, R. W., Reid F.M.H., Strickland J.D.H. 1970. Estimates of phytoplankton crop size, growth rate and primary production. Bull. Scripps Inst. Oceanogr. 17.
Grasshoff, K., Ehrhardt, M., Kremling, K. 1983. Methods of seawater analysis, 2ª ed., Verlag Chemie, Weinheim, 419 pp.
Huskin, I., Anadón, R., Wood-Walker, R, Head, B., Haris, R.M., Robins, D. (en prep.). Grazing impact of mesozooplankton in the Atlantic at a biochemical provinces scale.
Ikeda, T. 1985. Metabolic rates of epipelagic marine zooplankton as a function of body mass and temperature. Mar. Biol., 85: 1-11.
Kirchman, D.L., K’nees, E., Hodson, R.E. 1985. Leucine incorporation and its potential as a measure of protein synthesis by bacteria in natural aquatic systems. Applied Environ. Microbiol. 49: 599-607.
Kishino, M., N., Takahashi, N., Okami, S. Ichimura. 1985. Estimation of the spectral absorption coefficients of phytoplankton in the sea. Bull. Mar.Sci. 37: 634-642.
López-Urrutia, A. y Acuña, J.L. (Sometido). Gut Throughput dynamics in the appendicularian Oikopleura dioica.
Mackas, D y Bohrer, R. 1976. Fluorescence analysis of zooplankton gut content and investigation of diel feeding patterns. J. Exp.Mar. Biol.Ecol., 25: 77-85
Madin, L.P. y Cetta, C.M. 1984. The use of gut fluorescence to estimate grazing by oceanic salps. J. Plankton Res., 6 (3): 475-492.
Mague, T.H., Friberg, E., Hughes, D.J., I Morris. 1980. Extracellular release of carbon by marine phytoplankton; a physiological approach. Limnol. Oceanogr. 25(2): 262-279.
Malone, T. C. 1980. Algal size. In: The Physiological Ecology of Phytoplankton. Morris, I. (ed.) . Blackwell. pp: 433-463
Marañón, E., Holligan, P. M., Varela, M., Mouriño, B., Bale, A.J. 1999. Basin scale variability of phytoplankton biomass, production and growth in the Atlantic Ocean. Deep-Sea Res. (en prensa).
Porter, K.G., Feig, Y.S. 1980. The use of DAPI for identifying and counting aquatic microflora. Limnol. Oceanogr. 25: 943-948.
Slawyk, G., Raimbault, P., García, N. 1998. Measuring gross uptake of 15N labeled nitrogen by marine phytoplankton without particulate matter collection: Evidence of low 15N losses to the dissolved organic nitrogen pool. Limnol. Oceanogr. 43: 1734-1739.
Smetacek, V. 1975. Die Sukzession des Phytoplankton in der westlichen Kieler Butch. Diss. Kiel
Smith, D.C., Azam, F. 1992. A simple economical method for measuring bacterial protein synthesis rates in seawater using 3H-leucine. Mar. Microb. Food Webs 6: 107-114.
Strathmann, R. R. 1967. Estimating the organic carbon content of phytoplankton from cell volume or plasma volume. Limnol. Oceanogr. 12: 411-418.
Unesco (1994). Protocols for the joint blobal ocen flux study (JGOFS) core measurements. IOC. Manual and Guides. 170 pp.
Uthermöhl, H. 1958. Zur Vervollkommnung der quantitative Phytoplankton Methodik. Mitt. Int. Verein. Limnol. 9: 1-38.
Varela, R.A., Figueiras, F., Agustí, S., B. Arbones. 1998. Determining the contribution of pigments and the non-algal fraction to total absorption: toward a global algorithm. Limnol. Oceanogr., 43(3):449-457.
JUSTIFICACIÓN DE LA NECESIDAD DE COORDINACIÓN
(Sólo en el caso de proyectos coordinados)
La valoración de la propuesta se realiza como un todo y por ello es importante que destaque la necesidad de coordinar la actividad de los diferentes grupos para alcanzar los objetivos previstos.
El papel a desempeñar por cada grupo ha de estar en perfecta consonancia con el Plan de Trabajo y el presupuesto detallado de la propuesta. De hecho, las ventajas derivadas de la coordinación han de quedar bien reflejadas en la presente solicitud.
La consecución de los objetivos planteados en el proyecto supone la cuantificación simultánea de numerosas variables, cuya medida requiere la participación de especialistas en diferentes disciplinas dentro de la oceanografía biológica. Esta aproximación multidisciplinar hace aconsejable una estructura de proyecto coordinado cuyo desarrollo implica aglutinar un número elevado de investigadores de diversas instituciones.
Desde el punto de vista del desarrollo formal del proyecto, se ha mantenido la estructura de subproyectos por instituciones, dadas las ventajas que ésta supone para la gestión económica del proyecto. Sin embargo, desde la perspectiva científica, el proyecto se organiza en tres módulos que fueron ya definidos en la sección relativa al plan de trabajo y que aparecen representados en el esquema que se muestra a continuación.
Aunque cada una de las instituciones participantes es responsable de uno de estos módulos, no todos los investigadores de un módulo pertenecen exclusivamente al mismo organismo. Los módulos definidos son:
a) Estructura de biomasa y composición de las comunidades planctónicas
b) Circulación de C y N a través del microplancton
c) Efecto del zooplancton sobre la circulación de C y N en el ecosistema pelágico.
Cada módulo está coordinado por un investigador. Estos tres investigadores junto con el coordinador del proyecto constituyen el comité de coordinación del proyecto, que tiene como responsabilidad principal garantizar la integración de la información recogida en cada uno de los módulos. Los miembros de este comité se reunirán al menos tres veces a lo largo del proyecto (véase esquema de planificación temporal).
El coordinador del proyecto se ocupará de que todos los datos obtenidos en el proyecto tanto por los investigadores del propio proyecto, como aquellos proporcionados por la componente británica, se encuentren disponibles en la página web del proyecto. Asimismo, el coordinador garantizará la efectiva interacción entre CIRCANA y el componente británico AMT, para lo cual se prevé mantener dos reuniones de planificación e integración de resultados a lo largo del periodo de vigencia del proyecto.
Estructura organizativa y personal participante
En la tabla siguiente se indica la estructura organizativa del proyecto, así como la relación de los investigadores, becarios y técnicos que participarán en el mismo.
|
Coordinador Emilio Fernández |
||||
|
|
||||
|
Estructura de biomasa Coordinador Manuel Varela |
Flujos de C y N por el microplancton Coordinador Emilio Marañón |
Papel del zooplancton Coordinador Ricardo Anadón |
||
|
|
||||
|
Investigadores |
Investigadores |
Investigadores |
||
|
Florentina Álvarez |
Ramiro Varela |
Florentina Álvarez |
||
|
Ángeles Cid |
Ángeles Cid |
José Luis Acuña |
||
|
Santiago Fraga |
Manuel Varela |
Ana Miranda |
||
|
Ana Miranda |
Antonio Bode |
|||
|
Maite Álvarez-Ossorio |
||||
|
Becarios |
Becarios |
Becarios |
||
|
Eva M. Teira |
Jesús Cabal |
|||
|
Beatriz Mouriño |
Iñaki Huskin |
|||
|
Silvia Torres |
Mario Quevedo |
|||
|
Alejandro Isla |
||||
|
Ángel López |
||||
|
Personal técnico |
Personal técnico |
Personal técnico |
||
|
Jorge Lorenzo |
María José Pazó |
Jorge A. Sostres |
||
|
Pilar Iglesias |
Cristina Panete |
Esteban Cabal |
||
|
Isabel González |
Begoña Castro |
Gerardo Fernández |
||
|
Gerardo Fernández |